Реплисомы оказались взаимно «прозрачны» при терминации репликации
Причиной высокой скорости терминации репликации у позвоночных оказалась «прозрачность» прохождения двух реплисом с противоположных цепей ДНК друг мимо друга. Установить это удалось молекулярным биологам из Гарвардской медицинской школы с помощью разработанной ими новой модельной системы. Исследование опубликовано в Nature.
Механизм терминации репликации, то есть завершения синтеза ДНК, до сих пор изучен довольно плохо, по крайней мере по сравнению с другими стадиями репликации. Это связано, прежде всего, с методическими сложностями. Дело в том, что напрямую синхронизировать терминацию репликации в разных клетках очень тяжело. А без синхронизации невозможно увидеть процесс на молекулярном уровне, — по крайней мере без использования одномолекулярных методов (которые плохо подходят в этом случае).
 Плазмида Col E1 под микроскопом. Изображение: J. Inselburg, PNAS, 1974
Плазмида Col E1 под микроскопом. Изображение: J. Inselburg, PNAS, 1974
Создать модельную систему терминации репликации удавалось до сих пор только у бактерий, дрожжей и у эукариатических вирусов (SV40). Однако до сих пор было не понятно, насколько точно эти данные воспроизводят то, что происходит во время работы собственной ДНК-репликазы высших эукариот.
Модельная система, которую разработали молекулярные биологи из Гарварда, представляет собой плазмиду, в которую встроен сайт посадки для 16 репрессоров из лактозного оперона (длина сайта составляет 490 нуклеотидов). Эти репрессоры выступали в роли механических зажимов: они позволяли временно остановить продвижение реплисом из двух противоположных репликационных вилок. Затем, при добавлении специального реагента (IPTG), репрессорные «зажимы» одновременно «спадали» с ДНК во всех клетках и репликация синхронно завершалась. Это позволяло отследить все стадии процесса терминации стандартными молекулярно-биологическими методами (радиоактивным электрофорезом в агарозе и иммунопреципитацией хроматина).
Получив такую систему, авторам удалось показать, что при столкновении репликационных вилок не происходит снижения скорости синтеза ДНК, как предполагали ранее некоторые модели (причиной этого назывались топологические затруднения). Движущиеся друг на друга реплисомы совершенно не «чувствуют» друг друга вплоть до синтеза последнего нуклеотида, даже в момент расхождения цепей матричной ДНК. Синтез завершается с присоединением лидирующей цепи одной репликационной вилки к отстающей цепи противоположной вилки. После этого реплисомы диссоциируют с ДНК. Однако геликазы (белки, распутывающие ДНК перед движущейся полимеразой) двух репликационных вилок остаются связанными с нуклеиновой кислотой вплоть до полного лигирования всех вновь синтезированных фрагментов и распутывания (декатенации) двух дочерних молекул ДНК.
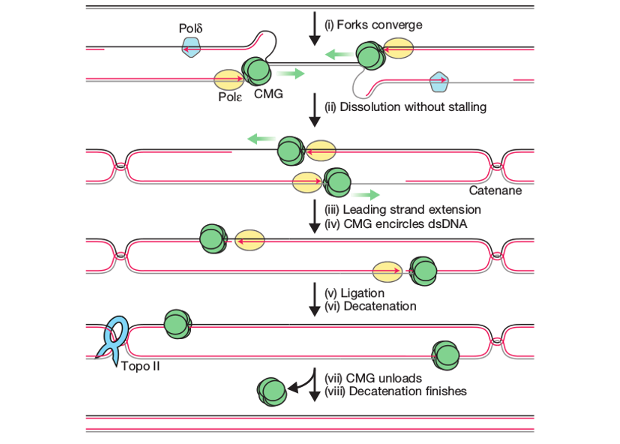 Последовательность разных стадий терминации репликации, установленная в новой работе. Изображение: James M. Dewar , Magda Budzowska, Johannes C. Walter, Nature, 2015
Последовательность разных стадий терминации репликации, установленная в новой работе. Изображение: James M. Dewar , Magda Budzowska, Johannes C. Walter, Nature, 2015
Наряду с транскрипцией и трансляцией репликация является одним из центральных процессов, которые исследует молекулярная биология начиная с 50-х годов прошлого века. Молекулярный механизм большинства из этих процессов установлен довольно давно и количество подобных «белых пятен» в базовых процессах на сегодняшний день очень невелико.
- Источник(и):
- Войдите на сайт для отправки комментариев
